
Introduction with a brief history of former studiesby Torsten Dikow & Fritz Geller-Grimm
The phylogenetic relationships of taxa within the monophyletic Asilidae are still very poorly known. The Asilidae attracted much attention from dipterists and many revisionary projects and ecological studies were undertaken. Despite this interest a comprehensive understanding of major evolutionary events during the phylogenetic history of the Asilidae are not known. This page will provide an overview of what is known so far and point out recent studies that incorporated morphological as well as DNA-sequence data to reconstruct the phylogeny of the Asilidae or subtaxa within it.
Recent studies
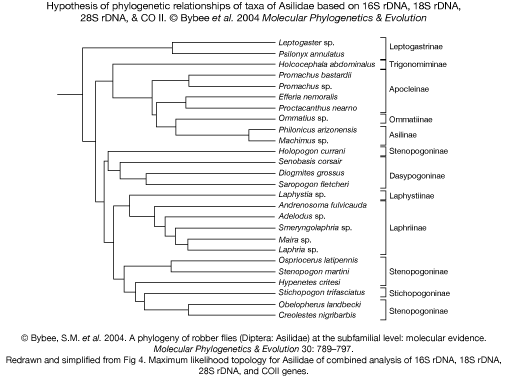
The first molecular hypothesis of the higher-level phylogenetic relationships within the Asilidae based on DNA-sequence data was recently published by Bybee et al. (A phylogeny of robber flies (Diptera: Asilidae) at the subfamilial level: molecular evidence. 2004. Molecular Phylogenetics & Evolution 30: 787797). The analysis is based on the 16S (mitochondrial), 18S, and 28S (both nuclear) ribosomal genes
and the protein coding gene Cytochrome Oxidase II (mitochondrial), sequenced from 26 representative species of 10 of the currently 11 recognised subfamilies. The results are preliminary, but this work provides the first molecular estimate of higher-level relationships. The following figure illustrates the major conclusions, which are: the
Leptogastrinae are basal; Trigonomiminae and Asilinae-Apocleinae-Ommatiinae form a monophylum; Laphriinae and
Laphystiinae form a monophylum; both Stenopogoninae and Dasypogoninae are paraphyletic.
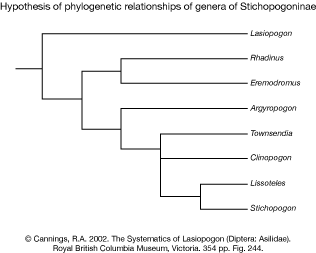
Cannings (2002) published a large revision of Lasiopogon and included a phylogenetic analysis of 8 genera of Stichopogoninae (not included were Afghanopogon and Stackelberginia). The hypothesis (see figure below) is based on 19 morphological characters from the complex male terminalia. He concluded that Lasiopogon is in fact a member of this monophyletic taxon and Stichopogon, the type genus, is a derived member of this group.
Papavero (1973)
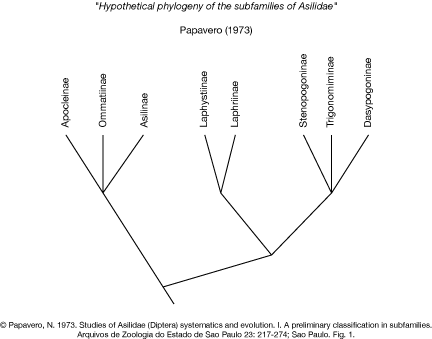
The most comprehensive morphological analysis was published by Papavero (1973). This analysis is based on a selected number of characters of the body of the imagines. The hypothesis of relationships of subfamilies of Asilidae is not based on the theory of phylogenetic systematics and therefore does not provide a modern hypothesis testable with these methods.
Papavero (1973a) stated: "For the establishment of the phylogenetic trends of the several groups of Asilidae, the following characters were taken in consideration by past authors, and will be employed here."
The list of characters and their interpretation by Papavero is provided here:
- Antennal style: The third antennal segment bears in most of the genera an apical style, which may be double or single. Primitive condition is represented by a style with 2 microsegments; this style undergoes reduction in many groups, until it becomes totally absent, being represented by only a minute apical spine, normally placed inside a small pit; this spine, as a derived condition, can be displaced to the dorsal surface of the 3rd antennal segment. Several genera present intermediary stages between a one-segmented style and a partially fused style.
A reduction of the antennal style has occurred many times, in many groups, as a notorious tendency, it cannot be used as a diagnostic character for subfamilies, serving merely to show the several trends within the family.
- Condition of the prosternum: The primitive prosternum was complete, fused to the pronotum, forming a complete ring. Later, it became emarginated, and finally separated into a dorsal and a ventral plate. broadly isolated by membrane. Some groups have always maintained the primitive condition, other morphological characters showing the derived condition; some other groups show all the steps, from complete to dissociated. This character is of tremendous importance to establish the early branchings of a group.
- Chaetotaxy: There is a general tendency among robber flies to reduce the bristles or hairs, especially the tergal bristles. Of course, many intermediary forms exist. However, within the Asilidae, two tendencies can be noted:
(i) A group that seems to have lost (or reduced) very early in its evolution the tergal bristles;
(ii) A group that primitively kept those bristles, and afterwards had them reduced.
Of great importance is also the chaetotaxy of the pleura, especially of the meso- and metapleura; according to this character, three groups can be more or less segregated:
(i) A group without differentiated meso- and metapleural bristles;
(ii) A group that preserved strongly differentiated mesopleural bristles (secondarily they may be reduced);
(iii) A group that preserved the metapleural bristles (also these may be secondarily reduced).
- Pilosity of the post-scutellar ("metanotal") slopes: Pilosity on the post-scutellar slopes seems to have been acquired (as a derived character) independently by several groups. This is also a good character for the establishment of evolutionary trends.
- Pilosity of antennal style: Several genera show a more or less pilose antennal style. A unique character is shown by a group of genera related to Ommatius, where the arista-like style shows a series of filaments on the underside.
- Wing venation: Most authors have relied on wing venation for the classification of Asilidae. The same as other characters, derived conditions appeared several times in several groups, and this character must be used with care.
(i) Marginal cell - may be open (primitive condition), closed, or closed and petiolate (derived condition);
(ii) First and fourth posterior cells, and anal cells same above;
(iii) Normally there are 5 posterior cells (primitive condition), which may become reduced to 4;
(iv) R3 - present in some genera (primitive condition); normally represented by a mere stump vein (intermediary condition), and in most genera completely absent (derived condition). Sometimes, the field of R3 is "reactivated" (Hennig, 1954), and a crossvein similar to R3 appears, forming 3 submarginal cells in the wing (as in certain species of Pseudorus); this is also a derived condition, that seldom occurs in the Asilidae; most genera with 3 submarginal cells (R3 complete) actually show the primitive condition of this character.
- Palpus: The primitively two-segmented palpus becomes reduced to a single segment. This has also occurred several times in the course of evolution.
- Apical spur on front tibiae: This character is very probably derived and monophyletic, as all the steps in its transformation can still be traced. Genera presenting this character were segregated under one subfamily.
- Female terminalia: The primitive female terminalia have spines on the acanthophorites; reduction of spines, fusion of the two halves of the acanthophorites and their subsequent elongation and modification leads to the formation of the several types of ovipositor presented by the different groups. This character, like the prosternum, is extremely important to determine the primitive branchings of the family.
- Male genitalia: The primitive condition is shown in the male genitalia by the free, triangular, hypandrium, the more or less elongate, simple basistylus, and the dististylus placed apically on the basistylus, and the epandrial halves free. The male genitalia seems to evolve more rapidly than the female terminalia, and a wide variety of types exist. These will be discussed in the next papers.
- Immature stages and fossils: Immature stages of a very limited number of genera are known (see Hennig, 1952), and no comprehensive study of the larvae has yet been made, so larval characters can be used in systematics. As to the fossils, a list of papers describing Asilidae (and Leptogastridae) fossils is presented as an Appendix to this paper; fossils described up to the present are, with a few exceptions, few, fragmentary, badly described, and erroneously identified. Therefore, no definite data may be obtained from these sources.
- Geography: This is an extremely important factor in establishing classification and phylogenies, and one that has been rarely used. I have tried to conform my groupings, as much as possible, to logical geographical (past and present) patterns.
Papavero then proposed the following character polarisation: "Hypothetical Phylogenetic History of Subfamilies. I believe that the Asilidae, like most of other families of Diptera, had their origin during the Mesozoic, in the supercontinent of Pangaea. The ancestral Asilidae stock should have the following primitive characters:
- Two-segmented palpus;
- Two-segmented antennal style;
- Prosternum complete, fused to pronotum, forming a complete ring;
- Fore tibiae without apical spur;
- Wings with 3 submarginal cells (i.e., R3 complete), and all cells (marginal, posterior and anal) widely open;
- Tergal bristles present on all tergites;
- Mesopleura and metapleura with several strong bristles;
- Female terminalia with spines on acanthophorites;
- Male genitalia with free hypandrium, free epandrial halves, and dististylus apically set on basistylus.
The ancestral stock has probably occupied all the available ecological areas and radiated rapidly, soon becoming differentiated in several branches, which originated the present subfamilies. In this initial differentiation, two trends may be discerned:
I. One group characterised primitively by the open wing cells, the almost complete loss of pleural and tergal bristles, by the spinous acanthophorites, but which very soon had the prosternum dissociated (while in some other genera the prosternum retained the primitive condition):
- One that acquired a twisted spine or spur on the fore tibiae - the Dasypogoninae;
- A branch where the head underwent strong modifications; the females lost the spines on the acanthophorites; deep mediogenal grooves on the face were developed; in most genera the prosternum remained with the primitive condition - the Trigonomiminae;
- The third branch remained relatively unchanged, and did not originally acquire derived characters, as two groups above; the prosternum remained complete in a limited number of genera (the Stichopogonini), but became dissociated in many others; similarly to the Dasypogoninae, most forms kept the original spinous acanthophorites - this branch originated the Stenopogoninae.
II. One group that kept most of the primitive characters; meso- and metapleural bristles, complete prosternum, wing venation. Group II became differentiated in two subgroups and several branches:
A. One which primarily kept the mesopleural bristles, and where females soon lost the spinous condition of the acanthophorites; this branch originated:
- 1. The Laphriinae with closed marginal cell and always with mesopleural bristles;
- The Laphystiinae with originally a mesopleural bristle, that later became absent, and with R4 strongly recurrent and sigmoid; the marginal cell remained open in some genera, closed in others.
B. A subgroup without differentiated mesopleural bristles, but with a row of bristles on the metapleura, always with closed marginal cell and a single-segmented palpus; the greatest mosaic of characters is obtained in this subgroup, which became modified in 3 branches:
- One where the antennal style became feathered, and where females lost the spinous acanthophorites - Ommatiinae;
- A branch with bare post-scutellar slopes; the most primitive genera have maintained the spinous acanthophorites, or wings with 3 submarginal cells; in other genera, the female terminalia became modified and the wings had their submarginal cell reduced to 2, R3 represented by a stump vein, or absent; the prosternum remained complete or was dissociated - the Apocleinae;
- Finally, in the third branch, the post-scutellar slopes became pilose, the wings had their submarginal cells reduced to 2, and R3 disappeared completely; the most primitive genera maintained the spinous acanthophorites, and the other developed different types of ovipositor - this group originated - the Asilinae."
Wood (1981)
Wood (1981) in the Manual of Nearctic Diptera diagnosed the Asilidae and listed a number of apomorphic character states shared by all species of robber flies.
"The Asilidae as here recognized are based on several synapomorphies. By far the most distinctive of these is the feeding mechanism and predatory behavior of the adult. The labella and prementum are fused to a form a heavily sclerotized, tube-like proboscis enclosing the needle-like hypopharynx that is used to stab the prey and inject it with saliva. The sexes are not dimorphic in this respect, unlike the Tabanomorpha and Nematocera. The mystax, a row or group of stout bristles along the lower edge of the face, is also peculiar to the Asilidae. The Mydidae, which are probably the sister group of the Asilidae, have a fleshy, pad-like labella, the primitive situation in the Diptera. Both families share a well-developed face and a sunken vertex, but the Mydidae lack a mystax. The Asilidae have retained the primitive number of three ocelli, whereas the mydids have only one. The larvae of both asilids and mydids are rather similar, especially in having a broadened paddle-like or scoop-like maxilla. However, in the Asilidae, the palpus arises in a notch situated laterally on the maxilla, not at the apex as in the Mydidae; also, the mandible does not extend as far forward as the apex of the maxilla but is hidden in a concavity along the median edge of the maxilla.
The Leptogastrinae, in spite of their many distinctive features (autapomorphies), share with the rest of the Asilidae the synapomorphies mentioned in the previous paragraph, namely the modified proboscis and predatory behavior of the adult and the modifications of the larval maxilla and mandible. Although Martin (1968a) proposed that the group be accorded full family rank, Oldroyd (1969) argued that there were insufficient grounds for separating them from the Asilidae. Further to Oldroyd's arguments, some of the autapomorphic characters of the Leptogastrinae, such as absence of the alula and the pulvilli, absence of acanthophorites of the tenth abdominal segment of the female, and absence of larval mandibles, must be considered, in the context of other Asilomorpha and Tabanomorpha, to be derived characters. Most other features of the Leptogastrinae such as their slender shape, their inclination to capture resting prey, their peculiar egg-laying habits, their helicopter-like flight, and their propensity to inhabit grassy habitats are autapomorphic; these features indicate only the distinctness of the Leptogastrinae as a group, not their phylogenetic position relative to other Asilidae or to the asilomorph families.
The Asilidae have been classified into a varying number of subfamilies and tribes (reviewed by Papavero 1973). Hull (1962) recognized five subfamilies, Dasypogoninae, Leptogastrinae, Laphriinae, Megapodinae, and Asilinae. With the exception of the Megapodinae, which are not found north of Mexico, Martin and Wilcox (1965) adopted the same arrangement as did Hull (1962). More recently, Papavero (1973a, 1973b) recognized eight subfamilies of Asilidae, not including Leptogaster and its allies which he considered as a separate family, following Martin (1968a). Papavero split the Asilinae into three subfamilies, Apocleinae, Asilinae, and Ommatiinae. The basis for separation of the Apocleinae seems to be the absence of hair or bristles on the anatergite. Most species of Asilus have these hairs, but a few species, e.g. auriannulatus Hine and blantoni Bromley, lack them and would therefore have to be placed in the Apocleinae. Although there may be some phylogenetic basis for the inclusion of most of these genera into a single taxon, it would probably be more convenient to group them at the tribal level. The Asilinae in broad sense (sensu Hull 1962) is probably a monophyletic group as well as a useful and readily recognized taxon; this interpretation is therefore used here, including the tribes Apocleini and Ommatiini.
The Laphriinae also seem to form a monophyletic group, reasonable distinct from Dasypogoninae in the immature stages as well as in the adult, but they may be an offshoot of some branch of the Dasypogoninae, such as the Laphystiini (no Laphystiini are known from the immature stages).
The Dasypogoninae (sensu Hull 1962) are a diverse group and are probably paraphyletic. Papavero restricted the Dasypogoninae to those forms with a stout sigmoid spine on the fore tibia. However, Ablautus and Omniablautus Pritchard, which are otherwise quite similar, differ in the presence or absence of this spine; likewise some specimens of a species that lack the spine and thus ought to belong to Wilcoxia James appears otherwise conspecific with specimens of Cophura which possesses the spine; therefore the spine may be present or absent in the same or closely related species. Hull (1962) has also remarked on the close similarity of some species that have the spine and those which lack it that would therefore be placed by Papavero in his Stenopogoninae.
The European species of Asilus were divided by Loew (1849) into several subgenera that were later raised to genera by subsequent authors. These taxa are similar to each other and are not readily separated. Theodor (1976) found differences in the internal terminalia of both sexes of some of these Palaearctic species that corresponded with the subgeneric arrangements proposed by Loew (1849). However, attempts by North American workers such Hine (1909) to place the Nearctic species in these European genera have not been entirely successful because many species are intermediate. Martin and Wilcox (1965) also attempted to place the Nearctic species into European genera, but left 38 species in Asilus (sens. lat.), even though they believed that these species were not congeneric with the type species of Asilus. Working with the fauna of Mexico and the south-western United States, Martin (1975) erected six new genera (including four occurring in the United States) for some of the species unassigned except to Asilus (sens. lat.) by Martin and Wilcox (1965), and regrouped other species did not fit readily into any of the categories. Most of the genera proposed by Martin (1975) were separated only by characters of the male terminalia; good characters are not available to separate most of the females ..."
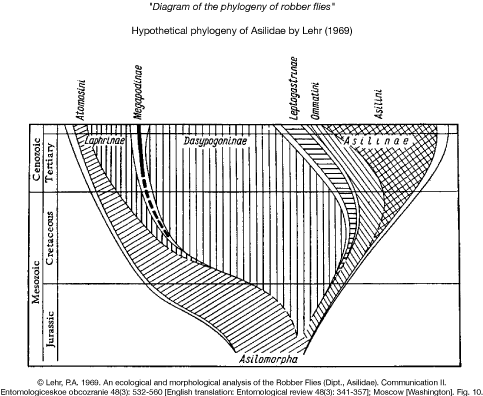
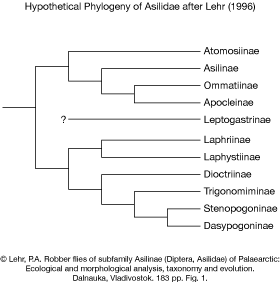
Lehr (1969 & 1996)
Lehr (1969 & 1996) published two different hypotheses of the phylogenetic relationships of Asilidae that are reproduced here.
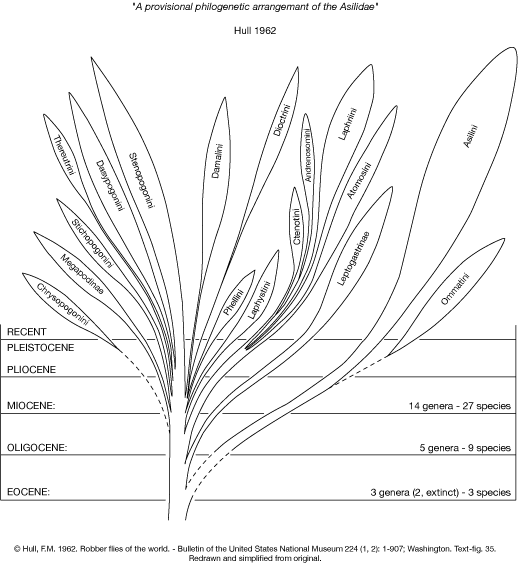
Hull (1962)
Hull (1962) published the following figure to illustrate his hypothesis of the phylogenetic relationships among taxa of Asilidae.
References
- Artigas, J.N. & Papavero, N. (1988): The American genera of Asilidae (Diptera): Keys for identification with an atlas of female spermatecae and other morphological details. I.Key to the subfamilies and subfamily Leptogastrinae. - Gayana. Zoologia. Universidad de Concepcion, Instituto Central de Biologia 52(1-2): 95-114; Concepcion.
- Artigas, J.N. & Papavero, N. (1988): The American genera of Asilidae (Diptera): Keys for identification with an atlas of female spermathecae and other morphological details. II. Key to the genera of Dasypogoninae Macquart, with descriptions of new genera and species and new synonymies. - Gayana. Zoologia. Universidad de Concepcion, Instituto Central de Biologia 52(3-4): 199-260; Concepcion.
- Artigas, J.N. & Papavero, N. (1991): The American genera of Asilidae (Diptera): Keys for identification with an atlas of female spermathecae and other morphological details. VII.1. Subfamily Stenopogoninae Hull. A preliminary classification into tribes. - Gayana. Zoologia. Universidad de Concepcion, Instituto Central de Biologia 55(2): 139-144; Concepcion.
- Artigas, J.N. & Papavero, N. & Pimentel, N.P. (1988): The American genera of Asilidae (Diptera): Keys for identification with an atlas of female spermathecae and other morphological details. IV. Key to the genera of Laphriinae Macquart (except tribe Atomosiini Hermann). - Boletim do Museu Paraense Emilio Goeldi. Serie Zoologia 4(2): 211-256; Belem.
- Bybee, S.M. & Taylor, S.D. & Nelson, C.R. & Whiting, M.F. [in press]: A phylogeny of robber flies (Diptera: Asilidae) at the subfamilial level: molecular evidence. Molecular Phylogenetics & Evolution.
- Cannings, R.A. (2002): The Systematics of Lasiopogon (Diptera: Asilidae). Royal British Columbia Museum, Victoria. 354 pp.
- Hull, F.M. (1962): Robber flies of the world. - Bulletin of the United States National Museum 224 (1, 2): 1-907; Washington.
- Lehr, P.A. (1969): An ecological and morphological analysis of the Robber Flies (Dipt., Asilidae). Communication II. - Entomologiceskoe obcozranie 48(3): 532-560 [English translation: Entomological review 48(3): 341-357]; Moscow [Washington].
- Lehr, P.A. (1996): Robber flies of subfamily Asilinae (Diptera, Asilidae) of Palaearctic: Ecological and morphological analysis, taxonomy and evolution. Dalnauka, Vladivostok. 183 pp.
- Papavero, N. (1973a): Studies of Asilidae (Diptera) systematics and evolution. I. A preliminary classification in subfamilies. - Arquivos de Zoologia do Estado de Sao Paulo 23: 217-274; Sao Paulo.
- Papavero, N. (1973b): Studies of Asilidae (Diptera) systematics and evolution. II. The tribes of Dasypogoninae. - Arquivos de Zoologia do Estado de Sao Paulo 23: 275-294; Sao Paulo.
- Wood, G.C. (1981): Asilidae. - In: McAlpine, J.F. & Peterson, B.V. & Shewell, G.E. & Teskey, H.J. & Vockeroth, J.R. & Wood, D.M. (Eds.): Manual of Nearctic Diptera. Volume 1. - Research Branch, Agriculture Canada, Monographs 27: : 549-573; Ottawa.
|